Bactrian Camels and Bactrian-Dromedary Hybrids
Daniel Potts1
University of Sydney
If the Silk Road may be described as ôthe bridge between Eastern and Western culures,ö then the Bactrian camel should rightfully be considered the principal means of locomotion across that bridge. Yet there is a great deal of misinformation concerning the Bactrian camel and its relatives, particularly in the ancient Near Eastern literature. This paper explores some of the problems surrounding Camelus bactrianus and the little-known hybrids of the Bactrian with the Arabian dromedary (Camelus dromedarius).
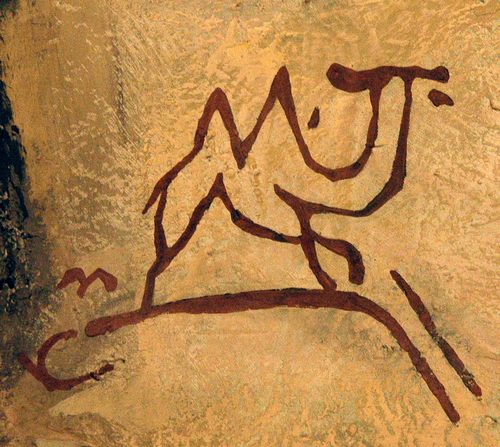
Fig. 1. Pre-historic cave image of camel. Display in
National Museum of Mongolian History, Ulaan Baatar.
Zoologists nowadays tends to favor the idea that Camelus bactrianus and dromedarius are descendants of two different sub-species of Camelus ferus (Peters and von den Driesch 1997: 652), and modern research suggests that the original habitat of the wild, two-humped camel extended from the great bend of the Yellow River in northwestern China through Mongolia to central Kazakhstan (Schaller 1998: 154; Nowak 1999: 1078; Bannikov 1976: 399) generally at elevations of 1500-2000 m. above sea-level. Although some scholars have suggested the original habitat of C. ferus may have extended as far west as the Caspian Sea, this is unlikely. If this were true, we should expect to find C. ferus faunal remains at prehistoric and early historic sites around the Caspian, but this is not the case. Moreover, to suggest that the natural distribution areas of the wild two-humped camel extended so far to the west flies in the face of everything that is known about the physiology and environmental adaptations of C. bactrianus (see below).
The survival of C. ferus in Inner Asia was long suspected but no firm evidence was available until N.M. Przewalski killed and described several specimens in 1873 (Camelus ferus Przewalski 1878 [?]). C. ferus has been described as ôrelatively small, lithe, and slender-legged, with very narrow feet and a body that looks laterally compressedö (Schaller 1998: 152).2 C. ferus has ôlow, pointed, coneshaped humps - usually about half the size of those of the domestic camel in fair conditionö (Bannikov 1976: 398). Representations of camels in the rock art of Palaeolithic caves in eastern Mongolia, such as Chojt-Zenker Cave, show what are believed to be C. ferus (Peters and von den Driesch 1997: 653, 661).3 (Fig. 1) C. ferus were still hunted in the medieval era in the Khotan, Turfan, Tarim, Lob and Katak regions of Inner Asia, and in Mongolia (Roux 1959-60: 50-51), while 18th-century Chinese records attest to the presence of wild camels on the northern and western edges of the Chinese empire (Lehmann 1891: 99). Small numbers are present in the region to this day (Heptner, Nasimovic and Bannikov 1966: 85-94; Bannikov 1976; Schaller 1998: 151-162).
The wild range of C. ferus, in all likelihood, extended only as far west as central Kazakhstan. This is significant for a number of reasons but first and foremost because this means that the natural distribution of the wild, two-humped progenitor of what we know as the domesticated Bactrian camel would not have included Bactria (northern Afghanistan/southern Uzbekistan) at all. How, then, to explain the name ôBactrianö given to the domesticated two-humped camel?
The term ôBactrianö was first applied to two-humped camels by Aristotle, who wrote of ôthe two species of camel, Bactrian and Arabianö (Historia Animalium 2.1 [498b9]) and noted that ôThe Bactrian camel differs from the Arabian in having two humps as against the latterĺs oneö (Historia Animalium 2.1 [499a15-17]). Some scholars have suggested that the name ôBactrianö became associated with the two-humped camel because camel-breeding developed in Bactria after initial domestication in eastern Iran and/or southern Turkmenistan (Schuegraf and Terbuyken 2001: 1225), but, as indicated above, this is not supported by the faunal evidence and it seems more likely that the Bactrian camel was introduced into Bactria proper from further east, not the south (eastern Iran/Seistan) or the west (Turkmenistan). Thus, like many commodities one can think of Ś Brussels sprouts or India ink Ś C. bactrianus would seem to be a misnomer. We have no idea where Aristotle got the designation ôBactrianö for the domesticated, two-humped camel, but he seems to have been responsible for introducing a term into the literature which should never have been applied to a mammal that was almost certainly domesticated outside of the region with which it is popularly associated.
Pure-bred Bactrians stand 1.5-2.4 m high to the top of the humps and are normally 1.68-1.63 m long, with a mean weight of 460 kgs (Epstein 1969: 118). They have been known to carry loads of 220-270 kgs some 30-40 kms daily, or 80-100 kms if pulling a loaded cart (Walz 1954: 56).4 Bactrian camels, which can live to be 35-40 years old, are generally put to work at the age of four and can expect to have 20-25 years of productive work (Epstein 1969: 120). They are at their best in the dry cold of the winter and spring months in Inner Asia, when their thick coats provided them with ample warmth. Able to withstand extremes of heat and cold, Bactrians prefer temperatures below 21░ C but are capable of tolerating a 70░-broad range between winter lows and summer highs (Manefield and Tinson 2000: 38). Consistent heat, however, is intolerable for Bactrians and the caravans which once set out from China westwards across the Gobi desert always traveled in winter (Walz 1954: 55-56). Bactrians have been known to function at altitudes of up to 4000 m. above sea level, e.g. in the Pamirs (Gauthier-Pilters and Dagg 1981: 6). After a long journey they were typically rested for 1-2 weeks and were pastured for the summer months on the steppe, where they built up their fat reserves again in anticipation of further caravan crossings the following winter (Walz 1954: 56).
The precise chronology of the gradual westward spread of the Bactrian camel is difficult to determine, but the available evidence nonetheless suggests we are dealing with a ôsloping chronology,ö i.e. a progression from earlier finds in the east towards later finds in the west. The principal difficulty that arises in verifying this hypothesis is the relative paucity of well-studied and dated faunal assemblages from sites within the range of C. ferus. Camelid faunal remains are said to have been found (Olsen 1988: 21)5 at Neolithic sites near Baotou (Inner Mongolia) and Lake Barkhol (northeast Xinjiang), and although it is not certain that these are C. bactrianus as opposed to C. ferus, they were certainly two-humped. The likelihood that these were domestic Bactrians is considered strong given that the locales would have been difficult to reach without the use of the camel (Peters and von den Driesch 1997: 661). As Lehmann wrote in 1891 (p. 141; my translation): ôWithout the camel neither the icy steppes of Western Siberia nor the inner Asian plains were inhabitable; they would have remained until today an insurmountable obstacle to communication and would have made a nomadic existence impossible.ö
By the middle of the fourth millennium BCE C. bactrianus was probably present in southern Turkmenistan. This being the case, and assuming a more or less continuous distribution of C. bactrianus from central Kazakhstan to the west, the animal was probably already present in Bactria by this time as well.6 Other sites on the southern (Iranian) side of the Kopet Dagh, however, do not show evidence of C. bactrianus at this early date.7 More Turkmenian evidence of C. bactrianus dates to the first half of the third millennium BCE. This includes terracotta models of wheeled carts drawn by Bactrian camels found at Altyn-depe in contexts dating to the Namazga IV period (Kohl 1992: 186) as well as faunal remains from Shor-depe, Chong-depe and Hapuz-depe (Compagnoni and Tosi 1978: Table 3).8 By the late third and early second millennium BCE the Bactrian camel is attested in the iconography of copper stamp seals and figurines thought to be from Bactria.9
Looking much further west, an unprovenanced cylinder seal in Old Syrian style in the Walters Art Gallery on which a Bactrian camel is depicted has been dated stylistically to c. 1750-1700 BCE (Gordon 1939: Pl. 7.55; Collon 2000: Fig. 8), but whether the fact that it bears a rider should be read as an indication that Bactrians were being ridden (e.g. Pohl 1950: 252) is unclear. Gordon noted that the awkwardness of the camelĺs form on the Walters seal indicated that the seal-cutter was unfamiliar with Bactrian camels. We cannot say whether this seal reflects the presence of Bactrian camels in the Syro-Anatolian area in the early second millennium BCE, direct contacts between Syro-Anatolia and a region in which Bactrians were present (whether native or introduced), and/or indirect contacts between such regions via intermediaries like Elam or Assyria.
A possible indication of the northwestward spread of the Bactrian camel by or during the third millennium BCE may be provided by faunal remains in todayĺs Tatarstan and Ukraine,10 and there is evidence of the southward and eastward spread of C. bactrianus into Pakistani Baluchistan beginning in the early second millennium BCE.11 As we move ahead into the Iron Age, there is little persuasive evidence to demonstrate the presence of Bactrian camels in western Iran.12

Fig. 2
This brings us, chronologically speaking, to the Achaemenid period when the Bactrian delegation, illustrated on the Apadana reliefs at Persepolis (Fig. 2), is shown bringing Bactrian camels to the imperial capital, and a Bactrian camel appears on one of the small gold plaques from the Oxus Treasure (Curtis and Searight 2003: Fig. 6.50). Thereafter, depictions of Bactrian camels become increasingly common, e.g. the Sarmatian gold plaques at Filippovka near the Ural River on the Eurasian steppes north of the Caspian (Aruz et al. 2000: Figs. 68, 96, 98) of fifth/fourth century BCE date. A particularly clear depiction from the late fifth century BCE occurs on a red-figured squat lekythos (E 695) in the British Museum (Curtius 1928: Abb. 6). Some scholars suggest that the Greeks first came into contact with Bactrian camels in Asia Minor, to which region they had been brought as a result of Achaemenid expansion (Schauenburg 1962: 99). Indeed Herodotus says that camels carried provisions for the advancing Persians, marvelling that Xerxesĺ camel train was attacked by lions while marching between Acanthus and Therma, even though the lions ôhad never seen that beast before, nor had any experience of itö (7.125). We do not know whether these were dromedaries, like those used by Cyrus against Croesus of Lydia (Herodotus 1.80).13
According to Soviet research (cited in Peters and von den Driesch 1997: 662), Bactrian camels were present and probably eaten during the Hellenistic period in Choresmia, between the Aral Sea and the Amu Darya (Oxus) River. Finally, Tĺang period (seventh/eighth century) tomb figures of Bactrian camels from China, some of which stand more than half a meter tall, are shown heavily laden with cargo (Vollmer, Keall and Nagai-Berthrong 1983: 47, 66) confirming their use as pack animals at this time.
In summary, the evidence just reviewed attests to an ever-expanding zone in which C. bactrianus is attested archaeologically outside the presumed native habitat of C. ferus. Although the data are not as plentiful as one would like, there is a general sense in which we move from the earliest evidence in the east (Neolithic Inner Mongolia) towards the west, with evidence beginning to appear in Turkmenistan (mid-fourth millennium BCE), Margiana and Bactria (mid-third millennium BCE) and surrounding areas to the north (Andronovo, Tripolye contexts) and south (Pirak, in Pakistani Baluchistan) as we move into the later second and early first millennium BCE. Above all, the available evidence flatly contradicts the idea that the two-humped camel was first domesticated in Bactria, and then spread eastward to China. In fact, it was precisely the opposite.
It is against the background of the archaeological evidence just reviewed that we turn now to some important epigraphic evidence attesting to the presence of C. bactrianus in Assyria from the end of the second through the middle of the first millennium BCE.
In a badly preserved fragment of the annals of Assur-bel-kala (1074-1057 BCE) on a tablet from Assur, the Assyrian king says that he sent merchants to acquire female Bactrian camels, udrate (Heimpel 1980: 331). On the Kurkh stele, Shalmaneser III (858-824 BCE) says that he brought back seven Bactrian camels as part of the booty from a campaign against Gilzanu (Mitchell 2000: 188, n. 7 with refs.), now thought to have been in the area south of Lake Urmia in northwestern Iran (Zadok 2002: 142-143). These are indeed illustrated twice, on the Black Obelisk, excavated by Layard at Nimrud, where they occur in Band 1 (Bulliet 1975: Fig. 70), and on the bronze gate decoration (Band 7) from Balawat, ancient Imgur-Enlil, several kilometers northeast of Nimrud (Bulliet 1975: Fig. 71). Interestingly, Bactrian camels are also shown in Band 3 on the Black Obelisk as tribute from Musri (Egypt).
Roughly a century later the Iranian stele of Tiglath-Pileser III (744-727 BCE), the exact provenance of which is unfortunately unknown, itemizes a long list of rulers from whom tribute was exacted, including several in the Zagros region of northwestern Iran. There we read, ôAnd as for Iranzu of Mannaea, Dalta of Ellipi, the city rulers of Namri, of Singibutu (and) of all the eastern mountains - horses, mules, Bactrian camels, cattle (and) sheep I imposed upon them (as tribute) to be received annually in Assyriaö (Tadmor 1994: 109). (15) In his first Babylonian campaign, the army of Sennacherib (704-681 BCE) seized both Bactrian and dromedary camels in Merodach-Baladanĺs abandoned camp near Kish (Luckenbill 1924: 56, l. 7). Esarhaddon (680-669 BCE) campaigned against Patusharra to the east of Assyria, seizing Bactrian camels as booty.16
A debt-note from the reign of Esarhaddon, dating to 674 bc (Postgate 1976: 149, no. 38), shows us an Assyrian official named Dannaya putting two Bactrian camels at the disposal of three individuals. The text reads (Kwasman and Parpola 1991: no. 241):
Two double-humped camels belonging to Dannaya, at the disposal of Yahutu, Ilu-kenu-[usur], and Adad-aplu-[ddina]. They shall give the camels back on the first of [Marchesvan] (VIII). If they do not give them, they shall pay 6 minas of silver. Month Tishri (VII), 14th day, eponym year of Sarru-nuri. Witness Siqi-Issar. Witness Sulmu-sarri. Witness Adad-dan. [I]f they do not pay the silver, it will increase by 2 shekels per mina.
These sources raise a number of questions. First, what of the lands from which Bactrian camels were acquired? With the possible exception of Patusharra, which may have been located as far east as the Pamirs (thus Vallat 1993), all of the regions mentioned as sources of Bactrian camels were situated in western Iran, from modern day Iranian Azerbaijan southwards to western Luristan. Yet as our review of the available faunal and other archaeological evidence of C. bactrianus has shown, nothing suggests that these areas were even remotely close to the most westerly regions where Bactrian camels are known in iconography and the faunal record. Indeed, judging by the faunal inventory at sites on the Iranian Plateau, Turkmenistan seems to be the most westerly of those areas where Bactrian camel use could be said to have become common. On the contrary, the presence of C. dromedarius remains at Chalcolithic Tepe Ghabristan (period 4, c. 3700-3000 BCE) and early Iron Age Tepe Sagzabad (late second millennium BCE) shows that the dromedary was known on the Iranian Plateau from an early date. In view of this evidence, it could be suggested that the Bactrian camels demanded of Mannaea, Ellipi, Namri, Singibutu or Gilzanu would not have been native to those districts, but themselves imported from further east. Presumably, however, the Assyrians would not have demanded Bactrian camels of these regions if they had not seen them there. The question arises, therefore, why were west Iranian communities keeping Bactrian camels, and might this have something to do with their eventual use by the Assyrians? The answer, I suggest, lies in camel hybridization.
From the seventeenth century onward, a series of European travelers, anthropologists and veterinary scientists have amassed an important record of observations of the intentional cross-breeding of Bactrian and dromedary camels (Kolpakow 1935; Menges 1935; Tapper 1985). As with most hybridization, the aim in crossing camels has been to produce a ôbetterö camel, in this case a more robust individual, stronger as a pack animal. In general, the best first generation hybrids are the products of male Bactrians crossed with female dromedaries, although female Bactrians crossed with male dromedaries are also attested. In cultures which practised camel hybridization, the stud function of Bactrian males was paramount, while small numbers of Bactrian females were kept in order to maintain a supply of pure Bactrian males. Tapper provides evidence on this practice over a region extending from Afghanistan to Anatolia. For example, amongst the Shahsevan of Azerbaijan, he observed, ôOnly the wealthiest of Shahsevan keep Bactrians, both male and female, and solely for breeding purposes. Female Bactrians are rarely if ever bought or sold, though they may change hands as gifts or be demanded as part of a bride-wealth.ö (Tapper 1985: 59-60). In Central Asia, Kolpakow (1935: 619) found that 6-7 year old Bactrians were optimal for use as studs.
Although larger and stronger, hybrids look like dromedaries in that they have one hump, though this is normally not very symmetrical and often has a small indentation between 4 and 12 cm deep which divides the rear portion of the hump Ś often 2-3 times as large as the front Ś from the front part. Alternatively, the hump may end up looking quite flat, and has been compared to a flattened pyramid. Hybridization produces a large animal, which can stand 2.32 m high at the hump or 2.15 m high at the shoulder (Kolpakow 1935: 618, n. 5). The legs are long, the height of the camel often greater than its length, and the weight sometimes in the 900-950 kg range, though more often approaching an average of c. 650 kg (Kolpakow 1935: 620).
All of the sources confirm the greater strength and load-bearing abilities
of the hybrids and indeed references to hybrids able to carry 400-500 kgs,
roughly double that of a dromedary and more than double that of an ordinary
Bactrian, are not uncommon. It should not be a surprise then that the
sources are also consistent in recording the substantially higher price of
hybrids vs dromedaries (Tapper 1985: 57, 59).
There is a considerable body of evidence concerning subsequent generations
of hybrids crossed with pure-bred or other hybrid individuals, all of which
points to their bad temper, inadequate size and generally poor quality. For
this reason, hybrid males were usually castrated (Tapper 1985: 61). As
Tapper (1985: 63) notes, ôThe hybrids were not allowed to breed, as their
offspring would be vicious and dangerous.ö Statements to the effect that the
dromedary-Bactrian crosses were infertile (Gray 1972: 161) are incorrect,
and ample evidence demonstrates the contrary (Peters and von den Driesch
1997: 654). Already in the early nineteenth century Eduard Friedrich
Eversmann reported seeing fertile offspring in Turkestan (Hartmann 1869:
70).17
To sum up, Tapper (1985: 67) notes, ôin my experience, the main advantage of the hybrid over the purer species, to both nomadic and commercial users, is less its supposed versatility than its vastly greater size, strength and carrying capacity, its aesthetically pleasing appearance, and its correspondingly greater value, in both financial and ceremonial terms.ö In view of the evidence just reviewed, which extends from Anatolia and Syria in the west to Afghanistan in the east, we can safely say that small numbers of Bactrian camels have been kept, over the past 300-400 years, by groups which, in the main, raised dromedaries, for the purpose of producing hybrids of outstanding strength. Further, these hybrids were used specifically as caravan and draught animals. It can at least be suggested, therefore, that the reason why Assyrian kings seized Bactrian camels and demanded them as tribute from Iranian lands to the east of Assyria was to acquire studs and breeding females in order to practice the same sort of hybridization as just outlined, with a view to developing stronger pack animals for a variety of purposes (military, commercial, agricultural). While we have as yet no archaeological evidence of camel hybridization from the Assyrian period, we do have some from later periods in the Near East, which demonstrates that hybridization was practiced in antiquity.
In recent years, archaeo-zoologists have identified faunal evidence of camel hybrids at Mleiha in the United Arab Emirates, Troy in western Turkey, and Pella in Jordan. Chronologically, the earliest evidence dates to the Roman or Parthian period. In 1994, Dr. S.A. Jasim excavated an important cemetery near Mleiha, in the interior of Sharjah, United Arab Emirates, which contained the graves of at least 12 camels, most of which, judging from associated finds, date to the first two centuries CE (Jasim 1999). The faunal remains, studied by H.-P. Uerpmann, included three hybrids. Identification of these individuals as hybrids was based on the morphometric analysis of selected bones (axis, astragalus, first phalanx) as compared with dromedary and Bactrian material. In addition to this material, Uerpmann (1999: 111-113) has identified the phalanx of a probable hybrid from a Roman context at Troy. Finally, I. K÷hler-Rollefson (1989: 149) has identified possible hybrids amongst the faunal remains from early Islamic Pella, in Jordan. These are presumed to have been killed by an earthquake in 747.
Additional archaeological evidence includes camel figurines from Parthian contexts at Seleucia-on-the-Tigris which, as Bulliet (1975: Fig. 80) noted nearly 30 years ago, exhibit the small indentation in the hump characteristic of the hybrid.
Conclusion
The ethnographic and historic observations reviewed above leave one in no doubt about the benefits and geographically widespread evidence of hybridization, while the archaeological evidence from Mleiha and Troy confirms the existence of hybrids by the Roman or Parthian period. But this evidence alone does not sustain Bullietĺs (1990: 731; cf. 1975: 168; Peters and von den Driesch 1997: 654) assertion that Diodorus ôcontains the first recorded reference to cross breeding of the dromedary and the Bactrian camel.ö I would suggest that the Neo-Assyrian sources cited above Ś the inclusion of Bactrian camels in the list of livestock demanded as tribute by Tiglath-Pileser III and Esarhaddon, the receipt of Bactrian camels from Musri and Gilzanu as shown on the Balawat gates and the Black Obelisk, the loan of Bactrian camels by Dannaya, Sennacheribĺs capture of Bactrian camels in Merodach-Baladanĺs camp, and Assur-bel-kalaĺs damaged reference to udrate Ś all point to the presence of Bactrian camels in Babylonia and Assyria some 500-1000 years before Diodorus observed them. Furthermore, given what we know of the distribution of C. ferus and C. bactrianus, and of the presence of C. dromedarius on the Iranian Plateau at an early date, I would suggest that the Iranian groups, mainly Median, from whom the Assyrians sought Bactrian camels were already engaged in camel hybridization by the time the Assyrians became conscious of the practice. Whether the Syrian cylinder seal from the eighteenth century BCE allows us to push that date even further back in time is difficult to answer, but in view of the ever-increasing body of evidence for ties between Central Asia and Elam (Amiet 1986: 146-207), and between Elam and Assyria and Mari in the early second millennium BCE (Potts 1999: 166ff), it is entirely possible that this was the period in which the peoples of the Near East first became aware of C. bactrianus. Whatever the case may be, it is now clear that the Bactrian camel has little beyond its name in common with the region of Bactria, and that its origins lie much further east, on the high steppes of Inner Asia. That it came to play an important role further west, already by the Assyrian period if not earlier, seems clear. The raison dĺŕtre behind the Assyrian interest in the Bactrian camel, and behind its later occurrence as far west as Anatolia, lies in the breeding of Bactrian-dromedary hybrids, the strength of which was un-surpassed by any other domestic animal, apart from the elephant, in the ancient Near East.
About the Author
Daniel Potts was educated at Harvard (AB ĺ75, PhD ĺ80) and has taught at the Freie Universitńt Berlin (1981-1986), the Univ. of Copenhagen (1980-1981, 1986-1991) and the Univ. of Sydney (1991-present). He is a specialist in the archaeology of Iran, Mesopotamia and the Persian Gulf, and has published widely on these areas. He is the founding editor of Arabian Archaeology & Epigraphy, and is a Fellow of the Australian Academy of the Humanities. He may be contacted at: dan.potts@arts.usyd.edu.au.
References
- Amiet 1986
P. Amiet. LĺÔge des Úchanges inter-iraniens, 3500-1700 avant J.-C. Paris, 1986. Notes et documents des MusÚes de France 11. - Amschler 1939
J. W. Amschler. Tierreste der Ausgrabungen von der ôgrossen K÷nigshŘgelö Shah TepÚ, in Nord-Iran. Stockholm, 1939. Reports from the Scientific Expedition to the North-West Provinces of China under the Leadership of Dr. Sven Hedin ŚThe Sino-Swedish Expedition Ś Publication 9. - Aruz et al. 2000
J. Aruz, A. Farkas, A. Alekseev, and E. Korolkova, eds. The Golden Deer of Eurasia. New York and New Haven: Metropolitan Mus-eum of Art and Yale University Press, 2000. - Badam 1984
G. L. Badam. ôHolocene faunal material from India with special reference to domesticated animals.ö In: Animals and archaeology: 3. Early herders and their flocks. J. Clutton-Brock and C. Grigson, eds. British Achae-ological Reports Int. Ser. 202. Oxford, 1984, pp. 339-353. - Bannikov 1976
A. G. Bannikov. ôWild camels of the Gobi.ö Wildlife, 18 (1976): 398-403. - Baur et al. 1933
P. V. C. Baur, M. I. Rostovtzeff, and A. R. Bellinger, The Excavations at Dura-Europus...Preliminary Report of Fourth Season of Work, October 1930 - March 1931. New Haven: Yale University Press, 1933. - Boardman and Vollenweider 1978
J. Boardman and M.-L. Vollen-weider. Catalogue of the engraved gems and finger rings (Ashmolean Museum, Oxford) I. Greek and Etruscan. Oxford: Clarendon Press, 1978. - Brentjes 1960
B. Brentjes, B. ôDas Kamel im Alten Orient.ö Klio, 38 (1960): 23-52. - Bulliet 1975
R. W. Bulliet. The Camel and the Wheel. Cambridge: Harvard University Press, 1975. - Bulliet 1980
R. W. Bulliet. ôCamel ii. In Persian history and economy.ö Encyclopaedia Iranica, 4 (1980): 730-733. - Collon 2000
D. Collon. ôLĺanimal dans les Úchanges et les relations diplomatiques.ö In: Les animaux et les hommes dans le monde syro-mÚsopotamien aux Úpoques historiques. Lyon: Topoi Supplement 2 (2000): 125-140. - Compagnoni and Tosi 1978
B. Compagnoni and M. Tosi. ôThe camel: Its distribution and state of domestication in the Middle East during the third millennium B.C. in light of finds from Shahr-i Sokhta.ö In: Meadow, R.H. and Zeder, M.A., eds. Approaches to faunal analysis in the Middle East. Peabody Museum Bulletin 2. Cambridge, 1978, pp. 91-103. - Curtius 1928
L. Curtius. ôSardanapal.ö Jahrbuch des Deutschen Archńologischen Instituts, 43 (1928): 281-297. - Curtis and Searight 2003
J. Curtis and A. Searight. ôThe gold plaques of the Oxus Treasure.ö In: Culture through objects: Ancient Near Eastern studies in honour of P.R.S. Moorey. T. Potts, M. Roaf and D. Stein, eds. Oxford: Griffith Institute, 2003, pp. 219-240. - Douglas van Buren 1939
E. Douglas van Buren. The fauna of ancient Mesopotamia. Rome, 1939. Analecta Orientalia 18. - Duerst 1908
J. U. Duerst. ôAnimal remains from the excavation at Anau and the horse of Anau in its relation to the history and to the races of domesticated horses.ö In: R. Pumpelly, ed. Explorations in Turkestan, Expedition of 1904. Washington: Carnegie Institution, 1908, pp. 341-442. - During Caspers 1972
E. C. L. During Caspers. ôLa hachette trouÚe de la sÚpulture E de Khurab, dans le Balouchistan persan, examen retrospectif.ö Iranica Antiqua, 9 (1972): 60-64. - Epstein 1969
H. Epstein. Domestic animals of China. Farnham Royal: Commonwealth Agricultural Bureaux, 1969. - Gauthier-Pilters and Dagg 1981
H. Gauthier-Pilters and A. I. Dagg, The Camel: Its Evolution, Ecology, Behavior, and Relationship to Man. Chicago and London: Univ. of Chicago Press, 1981 - Geiger 1979
W. Geiger. Ostiranische Kultur im Altertum. Aalen: Scientia, 1979 (repr. 1882 ed.). - Ghirshman 1938
R. Ghirshman. Fouilles de Sialk, prŔs de Kashan, 1933, 1934, 1937. Paris: Geuthner, 1938. - Gordon 1939
C. H. Gordon. Western Asiatic seals in the Walters Art Gallery. Iraq, 6 (1939): 3-34. - Gray 1972
A. P. Gray. Mammalian hybrids, 2nd ed. Slough: Commonwealth Agricultural Bureaux, 1972. - Hancar 1956
F. Hancar. Das Pferd in prńhistorischer und frŘher historischer Zeit. Vienna, 1956. Wiener Beitrńge zur Kulturgeschichte und Linguistik 11. - Hartmann 1869
R. Hartmann. ôStudien zur Geschichte der Hausthiere I. Das Kameel.ö Zeitschrift fŘr Ethnologie, 1 (1869): 66-79. - Heimpel 1980
W. Heimpel. ôKamel.ö Reallexikon der Assyriologie 5 (1980): 330-332. - Heptner et al. 1966
V. G. Heptner, A. A. Nasimovic, and A. G. Bannikov. Die Sńugetiere der Sowjetunion 1. Paarhufer und Unpaarhufer. Jena: Gustav Fischer Verlag, 1966. - van Ingen 1939
W. van Ingen. Figurines from Seleucia on the Tigris. Ann Arbor and London: Univ. of Michigan and Oxford Univ. Press, 1939. - Jettmar 1967
K. Jettmar. Art of the Steppes. New York: Crown Publishers, 1967. - Kohl 1984
P. L. Kohl. Central Asia: Palaeolithic Beginnings to the Iron Age. Paris: ╔ditions Recherche sur les Civilisations, 1984. - Kohl 1992
P. L. Kohl. ôCentral Asia (Western Turkestan): Neolithic to the Early Iron Age.ö In: R. W. Ehrich, ed. Chronologies in Old World Archaeology, 3rd ed. Chicago and London: Univ. of Chicago Press, 1992, pp. 179-195. - K÷hler-Rollefson 1989
I. K÷hler-Rollefson. ôZoological analysis of camel skeletons.ö In: Pella of the Decapolis, Vol. 2. Wooster: College of Wooster, 1989, pp. 142-164. - Kolpakow 1935
V. N. Kolpakow. ô▄ber Kamelkreuzungen.ö Berliner Tierńrztliche Wochenschrift, 51 (1935): 617-622. - Kuhrt 1999
A. Kuhrt. ôThe exploitation of the camel in the Neo-Assyrian empire.ö In: Studies in ancient Egypt in honour of H.S. Smith. A. Leahy and J. Tait, eds. London: Occasional Publications of the Egypt Exploration Society, 1999, pp. 179-184. - Kwasman and Parpola 1991
T. Kwasman and S. Parpola. Legal transactions of the royal court of Nineveh, Part 1. Tiglath-Pileser III through Esarhaddon. Helsinki, 1991. State Archives of Assyria 6. - Lamberg-Karlovsky 1969
C. C. Lamberg-Karlovsky. ôFurther notes on the shaft-hole pick-axe from Khurab, Makran.ö Iran, 7 (1969): 163-168. - Lattimore 1929
O. Lattimore. The Desert Road to Turkestan. Boston: Little, Brown, 1929. - Lehmann 1891
O. Lehmann. ôDas Kamel, seine geographische Verbreitung und die Bedingungen seines Vorkommens.ö Zeitschrift fŘr wissenschaftliche Geographie 8 (1891): 93-141. - Luckenbill 1924
D. D. Luckenbill. The Annals of Sennacherib. Chicago, 1924. Oriental Institute Publication 2. - Manefield and Tinson 2000
G. W. Manefield and A. H. Tinson. Camels: A compendium. Sydney, 2000. T.G. Hungerford Vade Mecum Series for Domestic Animals Series C 22. - Mashkour 2002
M. Mashkour. ôChasse et Úlevage au nord du Plateau central iranien entre le NÚolithique et lĺ┬ge du Fer.ö PalÚorient, 28 (2002): 27-42. - Mashkour et al. 1999
M. Mashkour, M. Fontugne, and C. Hatte. ôInvestigations on the evolution of subsistence economy in the Qazvin Plain (Iran) from the Neolithic to the Iron Age.ö Antiquity, 73 (1999): 65-76. - Mashkour and Yaghmayi 1996
M. Mashkour and E. Yaghmayi. ôFaunal remains from Tappeh Hessar (Iran), results of the 1995 excavation.ö In: Atti del XIII Congresso, Vol. 1. Forli: Unione Internazionale delle scienze preistoriche e protostoriche, 1996, pp. 543-551. - Meadow 1993
R. H. Meadow. ôContinuity and change in the agriculture of the Greater Indus Valley.ö International Association for the Study of the Cultures of Central Asia Information Bulletin, 19 (1993): 63-77. - Menges 1935
K. Menges. ôDie W÷rter fŘr ĹKamelĺ und einige seiner Kreuzungsformen im TŘrkischen.ö Ungarische JahrbŘcher, 15 (1935): 517-528. - Mitchell 2000
T. C. Mitchell. ôCamels in the Assyrian bas-reliefs.ö Iraq, 62 (2000): 187-194. - Nikylina 1994
N. M. Nikylina. Iskusstvo Ionii i Achemenidskogo Irana. [Art of Ionia and Achaemenid Iran.] Moscow: Iskusstvo, 1994. - Nowak 1999
R. M. Nowak. Walkerĺs mammals of the world, 6th ed. Baltimore: Johns Hopkins Univ. Press, 1999. - Olsen 1988
S. J. Olsen. ôThe camel in ancient China and an osteology of the camel.ö Proceedings of the Academy of Natural Sciences of Philadelphia, 140 (1988): 18-58. - Peters and von den Driesch 1997
J. Peters and A. von den Driesch. ôThe two-humped camel (Camelus bactrianus): New light on its distribution, management and medical treatment in the past.ö Journal of Zoology, 242 (1997): 651-679. - Pohl 1950
A. Pohl. ôDas Kamel in Meso-potamien.ö Orientalia, 19 (1950): 252. - Postgate 1976
J. N. Postgate. Fifty Neo-Assyrian legal documents. Warminster: Aris & Phillips, 1976. - Potts 1999
D. T. Potts. The archaeology of Elam: Formation and transformation of an ancient Iranian state. Cambridge: Cambridge Univ. Press, 1999. - Roux 1959-60
J.-P. Roux. ôLe chameau en Asie centrale.ö Central Asiatic Journal, 5 (1959-1960): 35-76. - Santoni 1979
M. Santoni. ôLes objets.ö In: J.-F. Jarrige and M. Santoni. Fouilles de Pirak, vol. 1. Paris: Boccard, 1979, pp. 167-232. - Sarianidi 1998a
V. Sarianidi. Myths of ancient Bactria and Margiana on its (sic) seals and amulets. Moscow: Pentagraphic, 1988. - Sarianidi 1998b
V. Sarianidi. Margiana and Protozoroastrism. Athens: Kapon Editions, 1988. - Schaller 1998
G. B. Schaller. Wildlife of the Tibetan Steppe. Chicago and London: University of Chicago Press, 1998. - Schauenberg 1962
K. Schauenberg. ôNeue antike Cameliden.ö Bonner JahrbŘcher, 162 (1962): 98-106. - Schuegraf and Terbuyken 2001
O. Schuegraf and P. Terbuyken. ôKamel.ö Reallexikon fŘr Antike und Christentum, 19 (2001): 1224-1241. - Shafer 1950
E. H. Shafer. ôThe camel in China down to the Mongol dynasty.ö Sinologica, 2 (1950): 165-194, 263-290. - Tadmor 1995
H. Tadmor. The inscriptions of Tiglath-Pileser III, king of Assyria. Jerusalem: Israel Academy of Sciences and Humanities, 1994. - Tapper 1985
R. Tapper. ôOne hump or two? Hybrid camels and pastoral cultures.ö Production pastorale et sociÚtÚ, 16 (1985): 55-69. - Uerpmann 1999
H.-P. Uerpmann. ôCamel and horse skeletons from proto-historic graves at Mleiha in the Emirate of Sharjah (U.A.E.).ö Arabian Archaeology & Epigraphy 10 (1999): 102-118. - Vallat 1993
F. Vallat. Les noms gÚographiques des sources suso-Úlamites. Wiesbaden, 1993. TAVO Beiheft B 7/11 [= RÚpertoire GÚographique des Textes CunÚiformes 11]. - Voigt and Dyson 1992
M. M. Voigt and R. H. Dyson, Jr. ôThe chronology of Iran, ca. 8000-2000 B.C.ö In: R. W. Ehrich, ed. Chronologies in Old World Archaeology, 3rd ed. Chicago and London: Univ. of Chicago Press, 1992, pp. 122-178. - Vollmer et al. 1983
J. E. Vollmer, E. J. Keall, and E. Nagai-Berthrong. Silk Roads, China Ships. Toronto: Royal Ontario Museum, 1983. - Walz 1954
R. Walz. ôNeue Untersuchungen zum Domestikationsproblem der altweltlichen Cameliden.ö Zeitschrift der Deutschen Morgenlńndischen Gesellschaft, 104 (1954): 45-87. - Wapnish 1984
P. Wapnish. ôThe dromedary and Bactrian camel in Levantine historical settings: The evidence from Tell Jemmeh.ö In: Animals and archaeology: 3. Early herders and their flocks. J. Clutton-Brock and C. Grigson, eds. British Arch. Reports Int. Ser. 202. Oxford, 1984, pp. 171-200. - Winkelmann 1999
S. Winkelmann. ôVon Formen und Stilen Ś ein Versuch der Gliederung baktrischer Stempel- und Rollsiegel.ö Hallesche Beitrńge zur Orientwissenschaft, 28 (1999): 112-209. - Zadok 2002
R. Zadok. The ethno-linguistic character of northwestern Iran and Kurdistan in the Neo-Assyrian period. Jaffa: Archaeological Center Publications, 2002. - Zeuner 1955
F. E. Zeuner. ôThe identity of the camel on the Khurab pick.ö Iraq, 17 (1955): 162-163.
Notes
1. This is a modified version of a lecture delivered for the Inner
Asia/Silkroad Study Group (IASSG) and co-sponsored by the Silkroad
Foundation and the Stanford University Center for East Asian Studies, 21
October 2004. The paper was based on the authorĺs ôCamel hybridization and
the role of Camelus bactrianus in the Ancient Near East,ö Journal of the
Economic and Social History of the Orient, 47 (2004): 143-165. Readers
interested in finding out more about the topic are advised to consult the
2004 article which has an extensive bibliography as well.
2. This thinness has given rise to the Mongolian name for C. ferus, havtagai,
from havtag, meaning Ĺflatnessĺ (Bannikov 1976: 398).
3. C. ferus are also well-represented in the rock art of the Altai, Tul-Kun,
Tamurasche, Uryankhai, Turgai and Minusinsk (Suljek) regions between Inner
Asia and Siberia but these are difficult to date (Heptner, Nasimovic and
Bannikov 1966: 87-88, Abb. 26; Peters and Von den Driesch 1997: 653).
4. Epstein 1969: 120, puts the average load at 120-150 kgs.
5. Olsen did not give a written reference for this information, but referred
to the fact that the camelid remains had been ôexcavated by archaeologists
from the Museum of Inner Mongolia in Huhhotö (Olsen 1988: 21), which he
visited in the early 1980ĺs. Dr. Jianjun Mei (Needham Research Institute,
Cambridge) kindly looked for written confirmation of this information but
was unable to find any.
6. Camelid remains found at Anau-depe in period II contexts were identified
as Bactrian on the basis of comparisons with modern Bactrian camels (Duerst
1908; cf. Compagnoni and Tosi 1978: 96 and Table 3), an identification that
has recently been reconfirmed (Peters and von den Driesch 1997: 658).
7. Tepe Hissar has no camelid fauna at all (Mashkour and Yaghmayi 1996)
while at Zagheh and Tepe Ghabristan (Qabrestan) on the Qazvin plain (west of
Tehran), only C. dromedarius seems to have been present in the fourth
millennium BCE (Mashkour, Fontugne and Hatte 1999: 71 and Table 2; Mashkour
2002: Table 2). (Dr. Marjan Mashkour [CNRS, Paris] kindly advised me that
the quantity of camelid remains at these sites was very small and therefore
somewhat surprising. She agreed that caution is required in interpreting
this data.) The depiction of a Bactrian camel on a sherd from period III4 at
Tepe Sialk (Ghirshman 1938: Pl. 79, A2) near Kashan has been interpreted as
evidence of C. bactrianus on the Iranian Plateau in the late fourth
millennium BCE (Compagnoni and Tosi 1978: Table 3).
8. Further south and east, skeletal remains of Camel sp. Ś possibly but not
definitely Bactrian Ś have been found in strata assigned to Period II
(phases 5-7) at Shahr-i Sokhta in Iranian Seistan (Compagnoni and Tosi 1978:
92), datable to c. 2700-2200 BCE (Voigt and Dyson 1992: 152), most probably
to the earlier part of this period. That Seistan, noted for its aridity, is
certainly well outside the natural habitat of the C. ferus is suggested by
the absence of camel remains in earlier, period I levels at the site, and
the absence of camels among the nearly 1300 zoomorphic figurines found there
(Peters and von den Driesch 1997: 656).
Faunal remains of probable Bactrian camel dating to the second half of the
third millennium BCE (Namazga V period) have been found at Ulug-depe,
Altyn-depe and Namazga-depe in southern Turkmenistan (Peters and von den
Driesch 1997: 659), where E.E. Kuzmina thinks ôthe use of Bactrian camels
for draft purposes was a local development...peculiar to southern Central
Asiaö (Kohl 1984: 114), and at Shah Tepe in northeastern Iran (Compagnoni
and Tosi 1978: Table 3; Amschler 1939: 77-80; but queried by Peters and von
den Driesch 1997: 660).
9. Their exact provenance is not known, since they were purchased on the
antiquities market (e.g. Amiet 1986: Fig. 189c; Sarianidi 1998a: 71-73, nos.
108-111; Winkelmann 1999: Abb. 4). Bactrian camel images are found on
soft-stone stamp seals from controlled excavations at Togolok 1 (burial 10),
Togolok 21 (southern court) and Gonur South (room 592) in Margiana (Sarianidi
1998a: 297, nos. 1634-1635). Gonur North has also yielded a theriomorphic
ceramic vessel in the shape of a Bactrian camel and a grafitto of a Bactrian
camel incised on a ceramic drain pipe (Sarianidi 1998b: Figs. 14-15).
Unfortunately, the camelid faunal remains from Gonur cannot be identified
to the species level (Meadow 1993: 72, n. 2). A clay bulla with a cylinder
seal impression from the temenos at Togolok 1 shows a human and a caprid
between two Bactrian camels (Sarianidi 1998b: Fig. 28.1 = Winkelmann 1999:
151 and Abb. 19.3).
10. Specifically, at the Andronovo site of Alekseevskoe in Tatarstan (Hancar
1956: 235; Heptner, Nasimovic and Bannikov 1966: 89 says C. ferus bones have
been found at Andronovo sites as well), in Karasuk culture graves at
Ilĺinskaia gora in the southwestern Ural foothills (Hancar 1956: Tab. 48 and
255; Brentjes 1960: 27), and possibly at the Tripolĺe culture site of
Gorodsk north of Kiev in Ukraine (Walz 1954: 79-80, n. 3; Hancar 1956: 69,
Tab. 22).
11. Camelid faunal remains from Harappan sites of late third and early
second millennium BCE have been identified in all cases as dromedary, not
Bactrian (e.g. Badam 1984: 349). However, a shaft-hole axe excavated by Sir
Aurel Stein in a grave at Khurab, in Iranian Baluchistan, is widely thought
to show a Bactrian camel in repose (Zeuner 1955; Lamberg-Karlovsky 1969;
During Caspers 1972). At Pirak in Pakistani Baluchistan the period IB-III
levels, dating to c. 1800-700 BCE, have yielded numerous clay figurines of
Bactrian camels (Santoni 1979: 177-179, Figs. 94-95, Pls. 42B and 43) as
well as faunal remains (Meadow 1993: 67, 70).
12. In Iran only C. dromedarius is present in the late second millennium BCE
levels at Tepe Sagzabad (Mashkour 2002: Table 2). We cannot be sure that the
so-called ôLuristan bronzeö buckle (4.7 cm. high, 8.7 cm. wide) in the
former Foroughi Collection in Tehran, dated stylistically to the 8th/7th
centuries BCE, on which a Bactrian camel is depicted (Jettmar 1967: Pl. 45)
is in fact a true reflection of the presence of Bactrian camels in western
Iran at that time. The provenance of the piece, even if it is authentic, is
simply too vague to attribute any geographical significance to it.
13. Bactrian camels appear on so-called ôGreco-Persianö seals of late
fifth/fourth century BCE date (Nikylina 1994: nos. 207, 216, 493) and on
Greek gems, probably of east Greek origin (Boardman and Vollenweider 1978:
114, no. 117). About this time, as well, the earliest references to Bactrian
camels occur in late fourth century BCE Chou documents in China, becoming
much more prevalent in the Han period (Shafer 1950: 174; cf. Walz 1954: 60;
Epstein 1969: 117). Brentjes has surveyed the terracottas and wall paintings
from the first millennium BCE and first millennium CE (Brentjes 1960: 28).
At least three fragments of Bactrian camel figurines were discovered by the
American excavators at Seleucia-on-the-Tigris in the 1930ĺs (van Ingen 1939:
320, no. 1465a-c and Pl. 76.557). Further Bactrian camel figurines of
Seleucid and/or Parthian date are known from Nippur (Douglas van Buren 1939:
36 with refs.).
14. Lattimore (1929: 133) observed, ôthe Mongols prefer a fast camel to a
pony for a long journey in haste, if they cannot get relays on the road,ö
favoring young camels which ôare much the best for riding, as they are not
only faster but softer-gaitedö (cf. Geiger 1979 [1882]: 456, on the use of
Bactrians by couriers in the Turkmenian deserts during the 19th century).
15. According to Zadokĺs (2002: 18ff, 42-3, 70, 77) recent study of western
Iran in the Neo-Assyrian period, Mannaea was located to the south of Lake
Urmia, Ellipi and Namri in western Luristan, and Singibutu around Khoy in
Iranian Azerbaijan.
16. He describes his campaign against Patusharra (Vallat 1993: 214-215), Ĺa
land that borders on the Salt Desert, in the midst of the land of the
distant Medes, near Bikni, the lapis lazuli-mountain, where none of my
forefathers had set footĺ. Patusharraĺs location is uncertain, and depends
on that of Mt. Bikni, a mountain which has been variously sought at Mt.
Alvand in Luristan, Mt. Demevend near Tehran, or in the Pamirs (for refs.
see Zadok 2002: 55).
17. The original source here is Eversmannĺs Reisen von Orenburg nach Buchara...nebst
einem naturhistorischen Anhange und einer Vorrede von H. Lichtenstein,
Berlin, 1823: 91.
